
Свою исследовательскую работу мы хотим начать с упоминания о Ноевом ковчеге, где собралось «каждой твари по паре», но описание основывалось только на представителях животного мира, исключая растительный. Вообще растениям всегда уделялось недостаточно внимания, как со стороны древних ученых и философов, так и в наше время. Нужно переосмыслить статус растений, отказавшись от антропоцентричной (философское представление, согласно которой человек является вершиной всех событий, то есть центром вселенной) картины мира, чтобы расширить понятия рациональности и сознательности, которые у растений, по нашему мнению, имеются, но которые следует изучать, отказавшись от привычных трактовок этих терминов.
Итак, у растений нет мозга и нервных клеток, по сравнению с животными они кажутся бесчувственными. Однако биологам известно, что представители этой группы многоклеточных организмов получают информацию извне и обрабатывают ее, могут общаться друг с другом с помощью химических сигналов.
Таким образом, тема нашей исследовательской работы «Особенности нейробиологии растений на примере Венериной мухоловки и Мимозы стыдливой», актуальна
Работы Юлиус фон Сакса, С.С. Пятыгина и многих других исследователей показывают актуальность проблемы, где предметом изучения является особенности нейробиологии растений.
Объект: нейробиология растений.
Предмет: особенности нейробиологии Венериной мухоловки и Мимозы стыдливой.
Цель: изучить особенности нейробиологии растений на примере Венериной мухоловки и Мимозы стыдливой.
Задачи:
1. Изучить характеристику особенностей нейробиологии растений;
2. Рассмотреть потенциал действий у растений;
3. Провести эксперимент, рассматривающую особенность жизнедеятельности Венериной мухоловки;
4. Пронаблюдать за особенностями жизнедеятельности Мимозы стыдливой.
Гипотеза: мы предполагаем, что если изучить особенности нейробиологии растений, то можно пронаблюдать механизм работы потенциала действий у растений на примере Венериной Мухоловки и Мимозы Стыдливой.
Для реализации поставленной цели и задач мы использовали следующие методы: теоретические (анализ литературы, синтез, обобщение взглядов ученых на проблему, классификация, систематизация, сравнение), эмпирические (беседа), наблюдение, эксперимент.
1. Теоретические основы нейробиологии растений
1.1. Характеристика особенностей нейробиологии растений
Знаете ли вы, что такое растительная нейробиология? Непосвященному человеку ее описание может показаться удивительным – это наука, изучающая систему коммуникаций растений, их сенсорные системы и «поведение». Нейробиологи утверждают, что растения умеют слышать, нюхать, общаться и чуть ли не видеть, а также манипулировать другими растениями и даже животными. Эти непривычные утверждения опираются на эксперименты, проведенные в лабораториях по всему миру, десятки лет работы и публикации в серьезных научных изданиях.
Когда мы пытаемся понять работу организма и его реакции в ответ на внешние воздействия, мы обычно обращаем внимание на его органы. Но у растения нет парных или одиночных органов вроде глаз или легких. Поэтому они, в определенном смысле, лучше защищены – потеряв оба глаза, животное лишается способности видеть и адекватно реагировать на внешнюю среду, а у растения все «органы» представлены во множественном числе. Оно может потерять до 90 процентов всего своего организма и все равно выжить. Если бы у растений, которые почти не могут двигаться, были бы такие же «слабые места», как у животных, то любая гусеница представляла бы для них серьезную опасность [6].
Растительный организм обладает способностью к определенной ориентировке своих органов в пространстве. Реагируя на внешние воздействия, растения меняют ориентировку органов. Различают движения отдельных органов растения, связанные с ростом – ростовые и с изменениями в тургорном напряжении отдельных клеток и тканей – тургорные. Ростовые движения, в свою очередь, бывают двух типов: тропические движения, или тропизмы,- движения, вызванные односторонним воздействием какого-либо фактора внешней среды (света, силы земного притяжения и др.). Любой тропизм может быть положительным или отрицательным. В зависимости от фактора, вызывающего тропические движения, различают геотропизм, фототропизм, хемотропизм, тигмотропизм, гидротропизм [5].
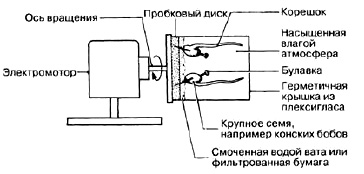
Геотропизм – движения, вызванные односторонним влиянием силы тяжести. Если положить проросток горизонтально, то через определенный промежуток времени корень изгибается вниз, а стебель – вверх. Еще в начале ХIХ в. был изобретен прибор клиностат (Приложение 1). В этом приборе проросток в горизонтальном положении привязывается к вращающейся оси. Благодаря этому сила притяжения действует попеременно на нижнюю и верхнюю стороны проростка. В этом случае рост проростка идет строго горизонтально и никаких изгибов не наблюдается. Эти опыты доказали, что изгибы стебля и корня связаны с односторонним действием силы земного притяжения [3].
Фототропизм – движения, вызванные неравномерным освещением разных сторон органа. Если свет падает с одной стороны, стебель изгибается по направлению к свету – положительный фототропизм. Корни обычно изгибаются в направлении от света – отрицательный фототропизм.
Гидротропизм – это изгибы, происходящие при неравномерном распределении воды.
Аэротропизм – ориентировка в пространстве, связанная с неравномерным распределением кислорода. Аэротропизм свойствен в основном корневым системам.
Тигмотропизм – реакция растений на одностороннее механическое воздействие. Тигмотропизм свойствен лазающим и вьющимся растениям.
Настические движения бывают двух типов: эпинастии – изгиб вниз и гипонастии – изгиб вверх. В зависимости от фактора, вызывающего те или иные настические движения, различают термонастии, фотонастии, никтинастии и др.
Термонастии – движения, вызванные сменой температуры.
Фотонастии – движения, вызванные сменой света и темноты. Цветки одних растений (соцветия одуванчика) закрываются при наступлении темноты и открываются на свету. Цветки других растений (табака) открываются с наступлением темноты. Механизм складывания листьев прост. Они поникают тогда, когда изгибается сочленение между черешком и листом. Изгиб этот, в свою очередь, происходит вследствие изменения давления клеточного сока, вызванного сжатием тела клетки под действием осмотически активных веществ – растворов солей, сахаров и других. При этом вода, находящаяся в клетках, перемещается в межклеточное пространство, вызывая падение давления клеточного сока. От того, в какой части сочленения происходит изменение давления сока, зависит поднятие или опускание листа [5].
Сейсмонастии – движения, вызванные толчком или прикосновением, например движение листьев у венериной мухоловки или у стыдливой мимозы. В результате прикосновения листья мимозы опускаются, а листочки складываются. Реакция происходит чрезвычайно быстро, спустя всего 0,1 с. При этом раздражение распространяется со скоростью 40–50 см/с. Так всем известная хищная Венерина мухоловка способна закрывать ловушки и переваривать насекомых и даже слизней.
1.2. Потенциал действий у растений
Хотя у растений нет ничего похожего на центральную нервную систему, они способны в ответ на различные раздражители генерировать распространяющиеся по организму электрические сигналы, напоминающие нервные импульсы животных. Эти сигналы не несут специфической информации о характере раздражителя и служат, по-видимому, для общей мобилизации защитных сил организма.
Хорошо известно, что у животных координация работы частей организма и целенаправленное поведение обеспечиваются в первую очередь деятельностью нервной системы. В ответ на те или иные стимулы нервные клетки генерируют электрический сигнал – потенциал действия (action potential), быстро распространяющийся по отросткам нейронов и передающийся от одного нейрона к другому при помощи специальных сигнальных молекул – нейромедиаторов – в местах межнейронных контактов (синапсах).
Гораздо меньше известен тот факт, что потенциалы действия (ПД) есть и у высших растений. Впервые их обнаружил около века назад индийский ученый Джагадис Чандра Бозе (Jagadish Chandra Bose), изучавший растение с двигательными реакциями – Mimosa pudica, которую иногда называют «электромимозой». Впоследствии ПД были обнаружены и у других высших растений.
Но у растений нет нервной системы, которая могла бы «анализировать» внешние сигналы и «принимать решения» на основе такого анализа. Нет у растений и специализированных образований, аналогичных аксонам – длинным отросткам нервных клеток, предназначенным для быстрого проведения электрических сигналов.
Сила ПД, то есть амплитуда импульсов, у животных и растений сходна: от нескольких десятков до сотни милливольт.
Генерация ПД и у животных, и у растений происходит по принципу «все или ничего». При возбуждении ниже порогового импульс не возникает совсем, а когда порог возбуждения достигнут, генерируется сразу максимальный по силе импульс.
По сравнению с животными, у растений ПД «замедлены» на 3–4 порядка. У животных длительность самого ПД и рефрактерного периода измеряется миллисекундами, у растений – секундами и десятками секунд (рефрактерный период – время, в течение которого клетка, сгенерировавшая ПД, остается невосприимчивой к новым стимулам и не может в ответ на них сгенерировать новый ПД). Соответственно и скорость распространения ПД у растений на несколько порядков ниже, чем у животных.
У животных принцип «все или ничего» не лишает нейроны возможности передавать информацию о силе раздражителя. Хотя сами по себе все ПД одинаковы по силе, в ответ на более мощный раздражитель нейрон может выдать серию из множества ПД, быстро следующих один за другим – нечто вроде «пулеметной очереди». У растений, напротив, ПД обычно одиночные, лишь изредка наблюдаются растянутые во времени серии из нескольких импульсов.
Правда, недавно в литературе появились сообщения об обнаружении у некоторых растений ритмически повторяющихся ультрабыстрых ПД, скорость распространения которых сопоставима со скоростью распространения нервных импульсов. Но отмечается, что растениям, лишенным нервной системы (как кодирующего и декодирующего органа) просто незачем генерировать серии таких ультрабыстрых ПД, и поэтому данное открытие, скорее всего, представляет собой очередной артефакт (ошибку).
В основе генерации ПД у растений и животных лежит возникновение пассивных ионных потоков через ионные каналы клеточной мембраны. В нервной клетке в «спокойном состоянии» мебрана поляризована: наружная сторона заряжена положительно, внутренняя – отрицательно. При возникновении ПД в мембране открываются ионные каналы, пропускающие ионы натрия. Положительно заряженные ионы натрия устремляются внутрь клетки, что приводит к деполяризации мембраны. Деполяризованный участок мембраны стимулирует такую же деполяризацию соседних участков – так распространяется ПД.
У растений ПД – это тоже деполяризация мембраны, но, в отличие от животных, она возникает не за счет входящего тока ионов натрия, а за счет исходящего тока ионов хлора. Кроме того, в генерации ПД у растений (в отличие от животных) важную роль играет активная работа «протонного насоса», выкачивающего протоны (H+) из клетки. После деполяризации мембрана снова поляризуется за счет исходящего тока ионов калия – это справедливо как для животных, так и для растений.
Но с животными все более-менее понятно. В ответ на те или иные стимулы нейроны генерируют серии ПД, поступающие в центральную нервную систему. Эти ПД несут информацию о силе стимула, а также о его характере, поскольку разные нейроны реагируют на разные стимулы. В центральной нервной системе эта информация в той или иной степени интерпретируется и анализируется, и в конечном итоге генерируются новые ПД – управляющие команды органам тела, которые обеспечивают адекватную реакцию организма на полученный сигнал.
У растений цепь событий должна быть радикально короче, поскольку у них нет никаких структур, хотя бы отдаленно соответствующих по своему назначению центральной нервной системе. Нет у растений и специализированных рецепторных клеток, ответственных за различение внешних стимулов.
По мнению С.С. Пятыгина, растительный ПД может быть эффективным сигналом о стрессирующем факторе, если в процессе распространения по растению он сам ведет себя подобно стрессору, то есть имитирует его влияние.
Таким образом, несмотря на некоторые черты сходства с нервным импульсом, ПД у высших растений представляет собой во многом уникальный системный сигнал. Основная роль его, по-видимому, состоит в имитации неспецифического влияния потенциально значимых для жизнедеятельности факторов за пределами зоны раздражения.
Особенно явно особенности потенциал действия проявляются у Венериной мухоловки и Мимозы стыдливой.
2. Экспериментальная часть
2.1. Характеристика особенностей потенциала действий у Венериной мухоловки
Эксперимент проводился в домашних условиях с 27 октября 2018 г. по 1 марта 2019 г. Для проведения эксперимента нам понадобился образец растения, который мы приобрели во флористическом магазине г. Читы.
Растение является видом, культивируемым в декоративном садоводстве. Может выращиваться как комнатная культура, но наше наблюдение показало, что нормальное развитие растения в комнатных условиях затруднено из-за недостаточной влажности воздуха и слишком высокой температуры зимой. Наше растение стало вялым, зелено-желтого цвета. Начинающий процесс цветения оказался подавлен. Мы предполагаем, что данное явления является следствием зимней спячки, так как растение было приобретено в осенний период, сменяющийся зимним (Приложение 2).

Наш образец (Венерина мухоловка) – небольшое травянистое растение с розеткой из 4–7 листьев, которые растут из короткого подземного стебля. Стебель – луковицеобразный. В период приобретения листья имели размер от трёх до семи сантиметров, длинные листья-ловушки были уже сформированы после цветения (Приложение 2).
Венерина мухоловка ловит своих жертв (насекомых, паукообразных) с помощью специализированного ловчего аппарата, образованного из краевых частей листьев. На внутренней стороне пластинок ловчего аппарата имеются тоненькие чувствительные волоски (Приложение 2). Экспериментальным путём мы выяснили, что для захлопывания ловчего аппарата необходимо оказать механическое воздействие минимум на два волоска на листе с интервалом не более 20 секунд. Такая характерная реакция обеспечивает защиту от случайного захлопывания в ответ на падение объектов, не имеющих питательной ценности (капли дождя, мусор и т. д.). Более того, переваривание начинается как минимум после пятикратной стимуляции чувствительных волосков. В открытом состоянии доли ловушки выпуклые (изогнуты наружу), после закрывания доли образуют изгиб, формируя внутри полость, выход из которой закрыт «ресничками». Верхняя часть каждой стороны ловушки Венериной мухоловки окрашена антоцианинами, пигментами, придающими поверхности ловушки красную расцветку. Мы считаем, что эта раскраска – главная приманка для насекомых у этого растения. Также наблюдалось образование небольшого количества клейкого вещества, являющееся разновидностью белка.
Подобный механизм описывается как бистабильная система с быстрым переключением, однако в настоящее время детальный механизм захлопывания ловушки понят не до конца. При механическом раздражении чувствительных волосков происходит генерация потенциала действия (значительную роль в данном процессе играют ионы кальция). Затем потенциал действия распространяется по долям ловушки и стимулирует клетки долей и средней жилки между долями. Предполагается, что у венериной мухоловки существует порог концентрации ионов, преодоление которого позволяет ловушке реагировать на стимуляцию. После закрывания венерина мухоловка «подсчитывает» дополнительные раздражающие волоски стимулы до пяти, после чего начинает секретировать пищеварительные ферменты.
Мы выяснили, что данные видоизмененные листья следствие недостатка азота в почве таких, как болота, где находится ареал обитания Венериной Мухоловки. Пойманные насекомые служат источником азота, необходимого для синтеза жизненно необходимых белков.
10 ноября мы провели экспериментальную работу по выявлению характерных особенностей потенциала действий Венериной Мухоловки. Для данного эксперимента пищей послужила Оса Обыкновенная. По нашим наблюдениям после того как оса оказалась помещенной в ловушку, при искусственной стимуляции спусковых волосков, произошла подача сигнала ловушке захлопнутся. Но более благоприятными условиями является естественное кормление с использованием растением клейкого вещества белкового происхождения, которое выделяет, привлекающий насекомых, запах.
Необходимо отметить, что механизм захлопывания ловушки у Венериной мухоловки можно подразделить на четыре основные фазы: Первоначальное захлопывание. Фаза сжатия. Фаза запечатывания. Фаза повторного открытия. «Спусковые волоски» являются индикаторами для растения, которое по их колебаниям определяет, что потенциальная добыча оказалась в ловушке. Если прикосновение произошло сразу к двум волоскам или к одному два раза подряд в течение 30 секунд, ловушка захлопывается за десятую долю секунды. Если первоначальное захлопывание ловушки успешно, начинается фаза сжатия, которая длится около получаса. Во время нашего эксперимента не произошло моментального закрытия ловушки, так как насекомое оказалось неспособным застимулировать необходимое количество волосков. Далее в процессе нахождения насекомого внутри ловушки и последующего соприкосновения со «спусковыми волосками», мухоловка продолжала дальнейшее зажатие створки, чтобы удержать жертву внутри. Если бы насекомое оказалось достаточно мало, оно способно проскользнуть сквозь зубцы ловушки и спастись. Фаза сжатия не происходит, если захлопывание оказалась неудачным в поиске добыче. Это бывает, если при схлопывании створок насекомое сумело выбраться из ловушки или произошло колебания волосков, вызванное другими листами, каплями дождя, или любопытством человека. В этом случае ловушка начинает медленно раскрываться и полностью откроется через день-два. Отметим, что если такое прикосновение к волоскам происходит несколько раз, это может привести к почернению и смерти ловушки. В последующие разы проведения эксперимента оказалось, что скорость схлопывания у одной и той же ловушки значительно падает.
Таким образом, наша ловушка успешно схватила добычу, насекомое не убежало перед началом фазы сжатия, следовательно, начался этап запечатывания. Во время этой фазы наблюдалось, как зубцы ловушки выдвигаются вперед и наружу, таким образом, что перестают переплетаться между собой, как это наблюдалось в первоначальной фазе. В результате края долек мухоловки (лезвия) с обеих сторон плотно прижались друг к другу. Печать становится плотной и непроницаемой, началось выделение пищеварительной энзимы. Насекомое погружается в субстанцию и постепенно переваривается (Приложение 2).

В течение последующих 5–12 дней ловушка осталась закрытой на время пищеварения. В это время продолжались выделяться пищеварительные энзимы, растворяющие мягкие ткани насекомого. Выделись питательные вещества, содержащиеся в тканях насекомого в той форме, в которой их может впитать лист мухоловки. Необходимо отметить, что количество времени, необходимое для полного переваривания добычи, зависит от возраста насекомого, ловушки и температуры окружающей среды, а так же от размеров насекомого. Дольше длится переваривание более крупных объектов питания. Чем старше ловушка, тем медленнее выделяются пищеварительные энзимы, чем теплее погода, тем быстрее происходит переваривание. Для идеального «обеда» насекомое должно быть размером в одну третью размера ловушки. Если насекомое слишком большое или какая-то его часть свисает из ловушки, запечатывание не может быть плотным, как и произошло на нашем примере. Из-за крупных размеров насекомого ловушка погибла, она почернела, отмерла и отпала от растения, но прежде окончив финальную стадию переваривания.
По окончанию эксперимента наблюдался продукт конечного переваривания – хитиновый скелет Осы Обыкновенной.
Итак, данный эксперимент помог нам пронаблюдать особенность потенциала действий на примере Венериной Мухоловки.
2.2. Характеристика особенностей потенциала действий у Мимозы Стыдливой
Для проведения эксперимента нам понадобился образец растения, который мы решили культивировать из купленных нами семян в цветочном магазине. Период посадки семян октябрь 2018 г.
В момент прорастания растения мы пронаблюдали появление ростка с двоякоперистыми парными листьями. Максимальный рост на данный момент Мимозы стыдливой составляет 15 см, в природных условиях достигает от 70 реже – до 150 см (Приложение 3).


В процессе наблюдения мы выявили, что у растения имеются чувствительные листы, которые свертываются при прикосновении к ним. Это происходит благодаря тому, что у основания черешков расположены водные мембраны, а на листьях есть сенсорные участки, которые реагируют на давление. При контакте вода перемещается к этому месту, и под её тяжестью листья сворачиваются и опускаются (Приложение 3). В ходе исследования было обнаружено, что мимоза стыдливая имеет свойство «запоминать» особенности каждого контакта. При неоднократном прикосновении к растению, которое не представляло угрозы для жизнедеятельности Мимозы Стыдливой, не происходил типичный реакционный ответ на возбудитель.
При наблюдении, мы выяснили, что растение способно «уставать». При частом раздражении, способность к движениям на какое-то время теряется. Для подтверждения данного факта, мы провели эксперимент: в течение которого растение подвергалось периодическому возбуждению. Данное действие привело к адаптации растения по отношению к источнику возбудителя.
Во время теоретического изучения материала, мы узнали, что растение может различать тип прикосновения. Недавно австралийские ученые обнаружили, что мимоза стыдливая способна самообучаться. Согласно результатам исследования, опубликованного в журнале Oecologia, мимоза «запоминает» особенности каждого контакта. При повреждении корней человеком или животным, растение выделяет «коктейль» из ядовитых веществ, включающий метансульфоновую, молочную, пировиноградную кислоты и разные соединения серы, что нередко приводило к отравлению скота на пастбищах, но при этом контакт с металлом и другими предметами механизма выработки не запускал.
В результате биологи нашли еще одно концептуальное отличие мимозы стыдливой от прочих представителей фауны: если большинство растений, использующих для своей защиты яд, выделяют его из своей надземной части – то объект их изучения делает это с помощью своих корней, на которых расположены крохотные узелки. Более того: они не только производят яд, но и анализируют химический состав окружающей среды, «принимая решение» о потенциальной опасности.
Во время наблюдений, мы выдвинули гипотезу, что Мимоза Стыдливая при изменении уровня освещенности, связанное со сменой времени суток, совершается настические движения: листья реагируют также, как при реакции на возбудитель, т.е. сворачивают листовые пластинки.
Для достоверности мы провели эксперимент, при котором растение помещалось в светонепроницаемую среду. Оказалось, что движения повторялись с определенной периодичностью, даже в отсутствие внешних стимулов, как свет. Выяснилось, что растение совершает данные листовые движения, короче длины суток и составляет примерно 22–23 часа, что и привело к нашим ошибочным выводам, т.к. период закрытия листков совпадал с наступлением вечернего периода.
Необходимо отметить, что цветение мимозы происходит с появлением длинного цветоноса, на котором располагаются небольшие фиолетовые соцветия в виде шариков. Цветение длится с мая по сентябрь. Опыление происходит при помощи ветра или насекомых, после цветения образуются крючковидные изогнутые бобы.
Заключение
В последнее время мы все чаще сталкиваемся с новыми исследованиями в области изучения растений, направленными на изучение жизнедеятельности последних. В результате сложилось новая научная дисциплина, получившая название нейробиологии растений и исследующая то, каким образом растения анализируют внешние сигналы и, исходя из этого «принимают решение».
В первой главе мы рассмотрели теоретический материал об особенностях нейробиологии растений. Мы узнали, что растительный организм обладает способностью к определенной ориентировке своих органов в пространстве. Уточнили, что реагируя на внешние воздействия, растения меняют ориентировку органов. Раскрыли понятие потенциала действия (ПД) у высших растений. Опираясь на исследования Джагадис Чандра Бозе, мы сравнили особенности ПД у животных и растений.
Во второй главе в экспериментальной части дана характеристика особенностей потенциала действий у Венериной Мухоловки и Мимозы Стыдливой. Эксперимент проводился в домашних условиях с 27 октября 2018 г. по 1 марта 2019 г. Мы пронаблюдали, что несмотря на некоторые черты сходства с нервным импульсом, ПД у высших растений представляет собой во многом уникальный системный сигнал. Основная роль его, по-видимому, состоит в имитации неспецифического влияния потенциально значимых для жизнедеятельности факторов за пределами зоны раздражения, что и подтвердилось на наших предметах эксперимента.
Таким образом, гипотеза, что если изучить особенности нейробиологии растений, то можно пронаблюдать механизм работы потенциала действий у растений на примере Венериной Мухоловки и Мимозы Стыдливой, доказана.
Приложение 1
Клиностат для наблюдения за геотропизмом и фототропизмом растений

Принцип работы клиностата, с помощью которого Юлиус фон Сакс провел эксперимент по выявлению геотропизма растений
Приложение 2
Образец растения, который был приобретен во флористическом магазине

Начавшийся процесс цветения и пожелтение листьев

Тонкие чувствительные волоски ловчего аппарата

Этап запечатывания

Приложение 3
Длина растения на момент последнего замера

Типичная реакция на возбудитель

Библиографическая ссылка
Шагин П.С. ОСОБЕННОСТИ НЕЙРОБИОЛОГИИ РАСТЕНИЙ НА ПРИМЕРЕ ВЕНЕРИНОЙ МУХОЛОВКИ И МИМОЗЫ СТЫДЛИВОЙ // Международный школьный научный вестник. 2019. № 3-3. ;URL: https://school-herald.ru/ru/article/view?id=1093 (дата обращения: 21.07.2026).