
Введение. Изучение закономерностей приспособления организма к факторам окружающей среды является фундаментальной задачей медицины. Гипотермия является одним из важнейших факторов, воздействующих на организм человека [1,2].
Вместе с тем, клинические наблюдения с использованием функциональных методов исследования, только усугубляют расхождения во взглядах на роль холодового фактора. Поэтому важное значение имеют результаты экспериментальных морфологических исследований по изучению реакции тканей организма на гипотермию [3,4,5].
Цель исследования: изучить динамику изменения морфометрических и плоидометрических параметров ядер гепатоцитов после воздействия гипотермии.
Задачи исследования:
1. Изучить периметр и площадь ядер гепатоцитов в постгипотермальном периоде.
2. Изучить площадь сечения цитоплазмы и ядерно-цитоплазматическое соотношение гепатоцитов в постгипотермальном периоде.
3. Изучить индекс накопления ДНК (ИНДНК) гепатоцитов в постгипотермальном периоде.
4. Изучить гистограмму распределения ИНДНК гепатоцитов в постгипотермальном периоде.
Материалы и методы.
Исследование выполнено на 25 крысах линии Wistar. Гипотермию моделировали путем помещения животных, находящихся в индивидуальных клетках, в воду температурой 5 °С, при температуре окружающего воздуха 7 °С. Критерием прекращения воздействия служило достижение животными ректальной температуры 20-25 °С, что соответствовало глубокой степени гипотермии. Время экспозиции было индивидуальным и в среднем составляло 40 ± 5 мин.
Животные умерщвлялись сразу после прекращения охлаждения, через 2 суток, 5 суток и через 14 суток после прекращения охлаждения. Обезболивание и умерщвление животных проводили в соответствии с «Правилами проведения работ с использованием экспериментальных животных». Материал фиксировали в 10 % нейтральном формалине забуференном по Лилли. Проводку материала осуществляли по изопропиловому спирту, заливали в парафин. Гистологические срезы толщиной 4-7 мкм окрашивали гематоксилином и эозином, для определения плоидности после стандартной гистологической проводки депарафинизированные срезы толщиной 4 мкм окрашивали методом Фельгена с использованием холодного гидролиза в течении 1 часа 30 минут. Цифровые микрофотографии микропрепаратов получали с помощью микроскопа Nikon Eclipse E200 (Япония) и цифровой видеокамеры VIDI CAM (Россия).
Морфометрию и плоидометрию осуществляли в морфометрической программе UTHSCSA Image Tool 3.0 (разработанной в University of the Texas Health Science Center of San Antonio, Texas). При плоидометрическом исследовании высчитывали индекс накопления ДНК (ИНДНК), равный среднему взвешенному показателю ДНК исследуемых клеток. В целях изучения распределения популяций по ИНДНК строили гистограммы гетерогенности клонов клеток.
Статистическую обработку материала проводили при помощи статистического пакета Statistica 6.0.
Гепатоциты сразу после воздействия гипотермии
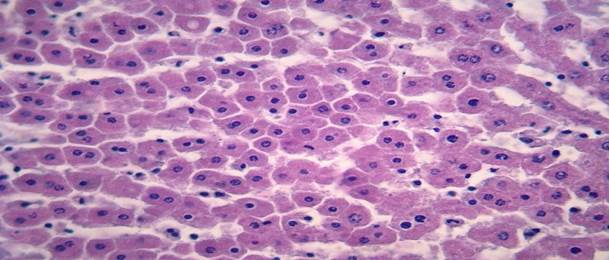
Рис. 1 Уменьшение размеров и пикноз ядер гепатоцитов печени крыс сразу после гипотермии. Окрашивание гематоксилином и эозином. Увеличение х 400.
Гепатоциты на 2 сутки после глубокой иммерсионной гипотермии
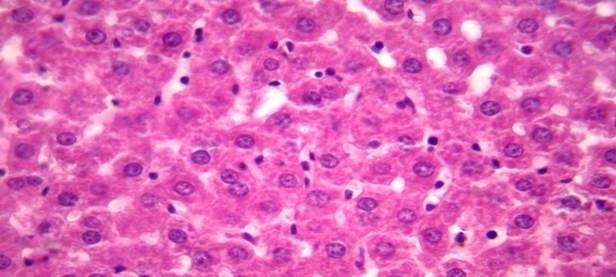
Рис. 2. На 2 сутки после глубокой гипотермии гепатоциты полиморфной формы. Ядра гепатоцитов увеличены в размерах. Окрашивание гематоксилином и эозином. Увеличение х 400.
Гепатоциты на 5 сутки после глубокой иммерсионной гипотермии
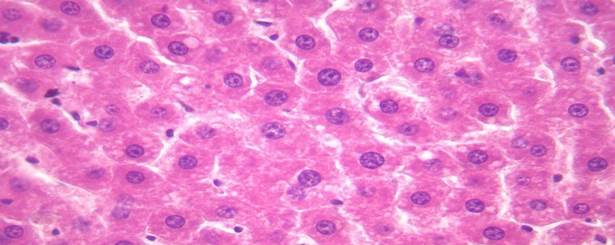
Рис. 3. На 5 сутки после глубокой гипотермии ядра гепатоцитов продолжают увеличиваться в размерах. Окрашивание гематоксилином и эозином. Увеличение х 400.
Гепатоциты на 14 сутки после глубокой иммерсионной гипотермии
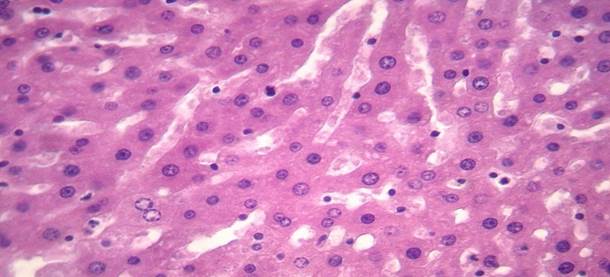
Рис. 4. На 14 сутки после глубокой гипотермии отмечается полиморфизм и уменьшение размеров ядра гепатоцитов. Окрашивание гематоксилином и эозином. Увеличение х 400.
Собственные результаты. Результаты проведенного исследования показали, что у животных забитых сразу после проведения гипотермии ядра гепатоцитов вокруг центральных вен были уменьшены в размерах, некоторые печеночные клетки находились в состоянии пикноза, в то время как в периферийных отделах печеночных балок ядра клеток были увеличены в размерах. Средняя площадь ядра составила – 16,6±0,7 мкм2, периметр – 19,4±1,7 мкм. Площадь сечения цитоплазмы составила 193,2±18,1 мкм2, ядерно-цитоплазматическое соотношение – 0,11±0,01. ИНДНК ядра составил 3,9с±0,2, пределы колебаний ИНДНК составили от 3с до 6,2с, при модальном значении 4,4с. Гистограмма распределения ИНДНК имела одиночный пик в области 4с и сдвиг влево.
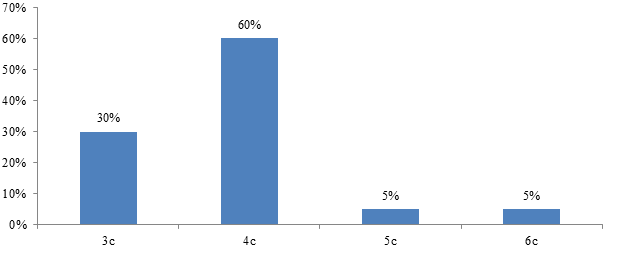 Рис. 5. Гистограмма распределения ИНДНК в ядрах гепатоцитов печени крыс сразу после проведения гипотермии. По оси абсцисс – плоидность гепатоцитов; по оси ординат – проценты.
Рис. 5. Гистограмма распределения ИНДНК в ядрах гепатоцитов печени крыс сразу после проведения гипотермии. По оси абсцисс – плоидность гепатоцитов; по оси ординат – проценты.
На 2 сутки после глубокой гипотермии ткань печени была в состоянии сильного отека, отдельные гепатоциты находились в состоянии некроза. Гепатоциты полиморфной формы. Средняя площадь ядра составила – 49,5±2,9 мкм2, периметр - 26,5±0,8 мкм. Площадь сечения цитоплазмы составила 260±7,1 мкм2, соотношение ядра и цитоплазмы – 0,19±0,01. ИНДНК составил 5,1с±0,2, пределы колебаний составили от 3,65с до 6,8с, при модальном значении 6с. Гистограмма распределения ИНДНК имела сдвиг вправо, наиболее высокий пик находился в области 6с.
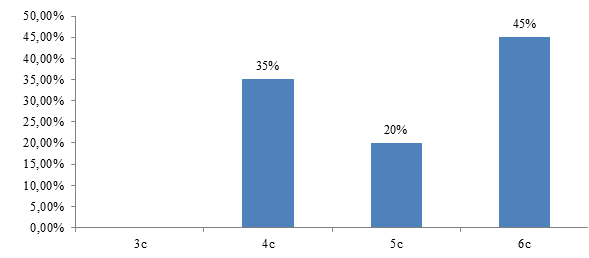
Рис. 6. Гистограмма распределения ИНДНК в ядрах гепатоцитов печени крыс через 2 дня после проведения гипотермии. По оси абсцисс – плоидность гепатоцитов; по оси ординат – проценты.
На 5 сутки после глубокой гипотермии ткань печени умеренно отечна. Гепатоциты полиморфной формы. Ядра гепатоцитов увеличены в размерах. Средняя площадь ядра составила – 72,7±4,5 мкм2, периметр – 32,1±0,9 мкм. Площадь сечения цитоплазмы составила – 436,4±34,9 мкм2, ядерно-цитоплазматическое соотношение – 0,17±0,01. ИНДНК составил 7,8с±0,4, пределы колебаний составили от 4,75с до 11с, при модальном значении 9,1с. Гистограмма распределения ИНДНК характеризовалась широкой гетерогенностью показателя ИНДНК, сдвигом вправо и наиболее высокий пик в был области 8с.
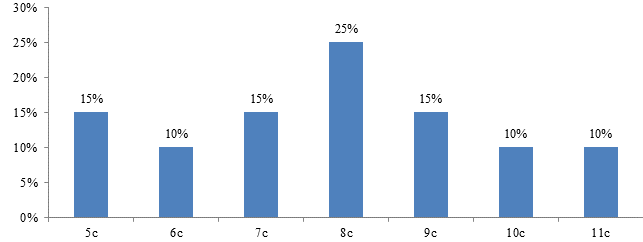
Рис. 7. Гистограмма распределения ИНДНК в ядрах гепатоцитов печени крыс через 5 дней после проведения гипотермии. По оси абсцисс – плоидность гепатоцитов; по оси ординат – проценты.
На 14 сутки после глубокой гипотермии ткань печени отмечали явления слабо выраженного отека. Ядра гепатоцитов по сравнению с предыдущем сроком эксперимента уменьшались в размерах. Средняя площадь ядра составила – 28,5±1,1 мкм2, периметр – 20,2±0,4 мкм. Площадь сечения цитоплазмы составила – 204,7±10,5 мкм2, соотношение ядра и цитоплазмы – 0,14±0,01. ИНДНК составил 4,1с±0,3, пределы колебаний составили от 2,4с до 4с, при модальном значении 3,9с. Гистограмма распределения ИНДНК характеризовалась сдвигом вправо, наиболее высокий пик отмечали в области 4с.
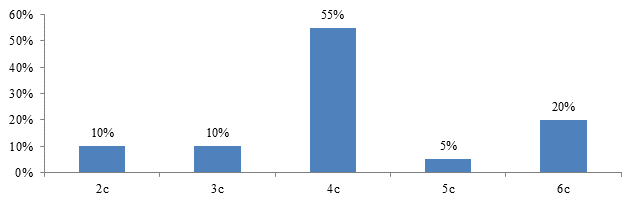
Рис. 8. Гистограмма распределения ИНДНК в ядрах гепатоцитов печени крыс через 14 дней после проведения гипотермии. По оси абсцисс – плоидность гепатоцитов; по оси ординат – проценты.
Заключение. Результаты проведенного исследования показали, что гипотермия приводит к уменьшению плоидности ядер и морфометрических параметров гепатоцитов у экспериментальных животных.
В отсроченные сроки после гипотермии, в период адаптации и регенерации клеток печени плоидность, морфометрические параметры ядра и цитоплазмы (площадь и периметр ядра, площадь цитоплазмы, ядерно-цитоплазматическое соотношение) увеличиваются.
К 14 дню после воздействия гипотермии морфометрические и плоидометрические параметры гепатоцитов приближаются к норме.